_fragment.jpg) |
The male flowers of Amborella are crowded with stamens.
Photo by Scott Zona, posted on Wikipedia |
The discovery that the lonely species
Amborella trichopoda occupies the tip of the most ancient known clade of flowering plants has caused quite a sensation in the botanical community. Here was the closest living relative of the common ancestor of all known flowering plants - a glimpse of what that ancestor might have been like! (Bailey & Swamy 1948).
Amborella is in the same position relative to angiosperms in genera, as
Acorus is to other monocots. I argued in a recent post (
What's so primitive about Acorus?), however, that being at the tip of a very long evolutionary branch does not necessarily mean being exactly the same as the very first members of that branch. It is likely in fact that some changes have been made as conditions and competition changed over the tens of millions of years since that ancient phylogenetic split. I identified several ways in which
Acorus was probably more advanced than some other archaic monocots that branched off slightly later.
 |
The female flower of Amborella
contains several carpels and a few
sterile stamens. The carpels are
ascidiate and unsealed. An opening
just below the stigma is blocked
only by a drop of fluid.
Photo by Sangtae Kim |
So we must consider that possibility with
Amborella. If
Amborella is the sister group to all other known angiosperms, it must have split off somewhere between 120 and 140 million years ago, certainly time for a few changes to have occurred. There may have been a whole family of amborellids early on, perhaps adapted for different light levels, soil types, pollinators, fruit dispersers, etc. Some may have even been flirting with the aquatic habitat, for the very next branch on the angiosperm tree consists of the waterlilies and their relatives. We have no idea - because at present at least, we have no fossil record of this lineage. All we have is the one species that survived to the present day.
To be sure,
Amborella trichopoda, has retained a number of truly archaic features. For one thing, it has the most primitive wood (consisting only of tracheids), of any living angiosperm (Carlquist & Schneider 2001). It also has very basic flowers, as we'll see below.
Amborella occurs in the rain forests of New Caledonian, a gentle environment isolated from both climate change and the hotbeds of aggressive evolution on the continents. Other relics of past ages survive in similar habitats, including the nearly-as-ancient order Austrobaileyales.
If we include the Austrobaileyales and the Nymphaeales with
Amborella in our analysis, we can create a more general picture of the ancestral angiosperm. Together these three clades are referred to as the ANITA grade, and contrast with the higher angiosperms of the magnolids, eudiots, and monocots. Each can be assumed to have a different mix of ancestral and specialized characteristics. The likely characteristics of the ancestral flower have been in fact derived from a study of this group (Endress 2001).
In this model, the ancestor of all known angiosperms had bisexual flowers consisting of simple, separate organs: tepals, stamens and carpels, which were spirally arranged and indefinite in number. Advancements from this model, such as carpels fused into a compound pistil, stamens in whorls of definite numbers, and tepals in two distinct whorls of sepals and petals, show up in various higher groups at different times.
Three other aspects of the carpels are also part of the model:
1. carpels are unsealed. They are open just below the stigmatic region, and entry of dirt, pathogens and small animals is blocked only by a drop of fluid. This contrasts with most modern carpels, which are completely sealed by a tight suture.
2. carpels are ascidiate, i.e. urn-shaped. The wall of the carpel is smooth and seamless, like a sock pulled up around its contents. This contrasts both with earlier accepted models of the first angiosperm carpels, and those of most modern angiosperms, which are plicate (folded). In the plicate model, a row of ovules along each margin of an ancient, leaf-like structure were brought inside as the margins joined together in a tight suture.
3. carpels contain just a few ovules placed opposite the backbone of the carpel, though there is some variation.
Amborella has just one ovule, some waterlilies have many.
 |
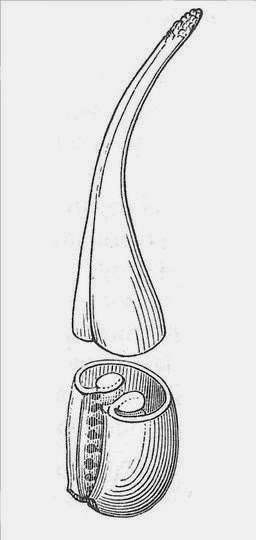
The carpel of Amborella is ascidate and unsealed at the top,
though the stigma region shows a folded structure
consistent with the plicate model of the carpel.
Drawing from Bailey & Swamy (1948) |
 |
In the plicate, or folded-leaf
model, the first carpels folded
or rolled together, bringing rows
of marginal ovules inside. The
two edges eventually became
tightly sealed by an interlocking
suture.
Drawing from the 1879
textbook by Asa Gray |
Assuming that this correctly describes the flower of the common ancestor,
Amborella is likely specialized in several ways. First the flowers are relatively small compared to others like most waterlilies,
Austrobaileya, and
Illicium, and massed together in an apparent group display. Masses of small, whitish flowers are common among eudicots, magnolids and monocots. Examples include viburnum, most palms, and carrots/Queen Anne's lace. Such displays are adaptation for a particular pollination strategy involving a variety of insects and possibly wind (Thien, et al. 2003).
The flowers are also unisexual, with pollen-producing flowers on separate plants from those that bear ovule-producing flowers. Such an arrangement is likely a means of avoiding self-pollination (Ferrandiz et al. 2010). Similar patterns can be seen in a variety of other plants, such as date palms. The fact that the female flowers contain sterile stamens between the tepals and the carpels is compelling evidence that the ancestors of
Amborella did indeed have bisexual flowers. This has recently been confirmed by Sauchet et al. (2017).
The fruits of
Amborella are described as small drupes. These are fleshy fruits with large seeds filled with food reserves, typically adapted for germination in shady forests. Drupes are found among many different families of flowering plants, but to my knowledge are always the endpoints of evolution from more generalized ancestors with flexible ovule production. In the Rose family, for example, drupes are found in the genus
Prunus (plums, cherries, etc.), a specialized genus in a family that includes a wide variety of fruit types.
 |
The bisexual flowers of Austrobaileya have a
number of carpels, each containing two rows of
ovules, as well as flattened, blade-like stamens.
Source: http://www.naturalist.if.ua/?p=3585 |
Drupes are constrained developmentally to produce only one ovule, which becomes surrounded by a pit, a hard layer the develops from the inner fruit wall. Reorganization of the development process to begin producing more ovules, and in two rows at that, would require numerous genetic changes. It would conceivably happen if there were selective pressure to produce more, smaller seeds, but this could be much more easily accomplished by increasing the number of carpels in the flower, or by increasing the number of flowers produced. Such a rigidly one-seeded carpel is therefore not likely the ancestor of the variety of carpels and fruit types we find among angiosperms today, or even what we find in the ANITA grade. Something like the multi-seeded fruit of
Austrobaileya is a much more likely starting point for all kinds of fruits, including drupes, berries, and capsules that open to release their seeds.
In sum,
Amborella is likely specialized in its floral display, unisexual flowers, and single-seeded carpels. In other ways, it does show its age: wood without vessels, simple, separate flower parts of indefinite numbers, and unsealed carpels. Its present very limited distribution in forests of New Caledonia attest to its archaic status and its proximity to extinction. The few ways in which it has specialized are probably the keys to it still being with us.
This discussion leads to a deeper question of the nature of the first angiosperm carpels, which evolved well before the common ancestor of living angiosperms. Were they ascidiate or plicate? The ascidiate carpel itself may be an adaptation for making berries and drupes, maybe nuts and achenes as well, but does not lend itself to carpels that must reopen as capsules, follicles or legumes. So were the very first carpels fleshy berries? I'll take that up in a future post.
References:
Bailey I. W. and Swamy B. G. L. 1948
Amborella trichopoda Baill., a new morphological type of vesselless dicotyledon. Journal of the Arnold Arboretum 29: 245–254.
Carlquist, S. J. and E. L. Schneider. 2001.
Amborella trichopoda: relationships with the Illiciales and implications for vessel origin. Pacific Science 55 (3): 305-312
Endress, P. K. 2001. The Flowers in Extant Basal Angiosperms and Inferences on Ancestral Flowers
International Journal of Plant Sciences 162 (5): 1111- 1140
Thien, L. B. , T. L. Sage, T. Jaffré, P. Bernhardt, V. Pontieri, P. H. Weston, D. Malloch, H. Azuma, S. W. Graham, M. A. McPherson, H. S. Rai, R. F. Sage and J-L. Dupre. 2003. The Population Structure and Floral Biology of
Amborella trichopoda (Amborellaceae) Annals of the Missouri Botanical Garden 90 (3): 466-490.
Sauquet, Hervé, Balthazar, Maria von …Schönenberger,Jürg. 2017 The ancestral flower of angiosperms and its early diversification. Nature Communications volume 8, Article number: 16047











