The carpel is the distinctive seed chamber of the angiosperms, or flowering plants. It is in fact the definitive feature of this major group. When the first carpel evolved, the first angiosperm came into existence. The carpel encloses, protects, and facilitates the fertilization of the ovules, which then mature as seeds. The carpel then becomes the fruit, and participates in the dispersal of the seeds. In the flower, carpels occupy the center and are surrounded by stamens and tepals (petals and sepals). In most modern angiosperms the carpels are joined together into a compound pistil (see "Were the first monocots syncarpous?)"
In those flowers in which the carpels remain separate, there are two fundamental shapes: plicate - resembling leaves that have folded with the opposite edges sealed together, and ascidiate - shaped like a vase or an urn.
 |
| The ascidiate carpels of Amborella contain a single seed, and have a large, folded stigma. A. the carpel at the time of pollination. B. the mature fruit, which is a drupe. Drawing from Bailey and Swamy, 1948. |
Above the opening, which is blocked only by a drop of fluid, the stigma is typically prolonged along what can be described as the backbone of the carpel, and ovules are attached in a line on the opposite side (i.e. in what might be called the "belly"), within the urn-shaped base. Sometimes, as in Amborella, the stigma is folded at the backbone, forming narrow flaps along each side. During the growth of the carpel, tissues at the base (“a meristematic cross-zone between the primordium margins” – Endress & Doyle 2009) push the wall upward around the ovules.
 |
| After pollination the carpels of Austrobaileya spread apart as they swell with the developing seeds within. Photo courtesy Dennis Stevenson. |
 |
| The carpels of Austrobaileya are ascidiate, but contain two rows of ovules opposite the backbone. The stigmas are pushed together to form a common head for receiving pollen. Photo courtesy Dennis Stevenson. |
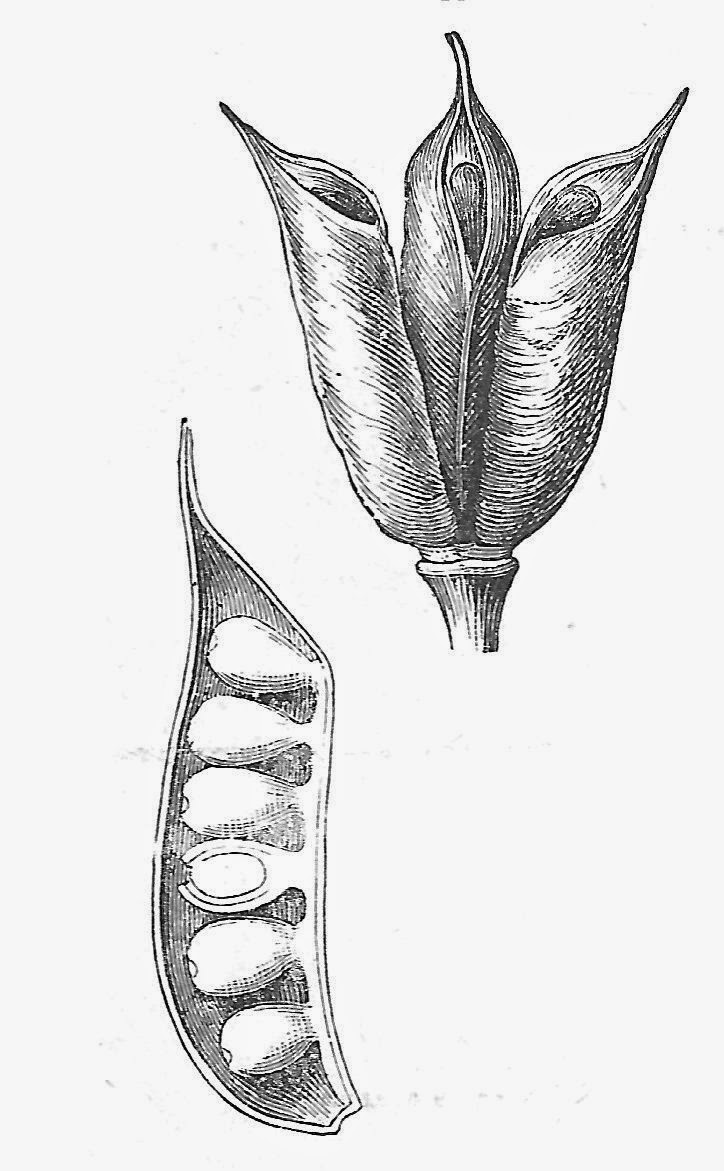 |
| Simple plicate carlpels, called follicles, can be seen most readily in the Ranunculaceae, in genera like Aquilegia (columbine), Delphinium, Eranthis, and others. Drawing from Asa Gray's Botanical Textbook, 1879. . |
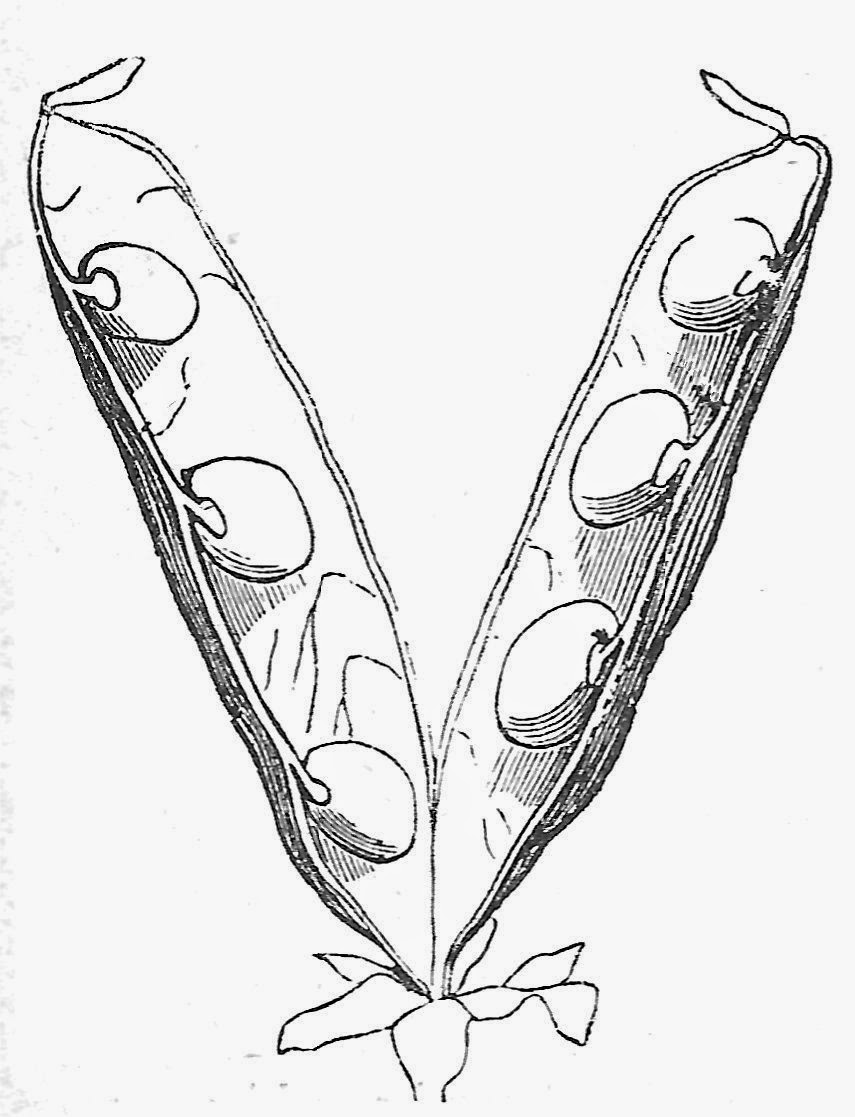 |
| The pea pod is somewhat more specialized than a follicle, as it has adapted to split along both the suture and the backbone when it opens to release its seeds. The seeds alternate along the two margins. Drawing from Thomé 1877, Textbook of Structural and Physiological Botany. |
 |
| Follicles of the genus Eranthis (Ranunculaceae) are quite leaf-like. |
As I have argued in earlier posts, however, these theoretical conclusions based on probability need to be tesed in the adaptive arena. In other worids, do they make sense in terms of "adaptive parsimony?" (see "Were the first monocots syncarpous?" for an explanation of this term)
 |
| In ancient seed ferns, such as this Sphenopteris, seeds were borne directly on large, frond- like leaves. Drawing from Brown, 1935, The Plant Kingdom. |
An interesting alternate idea is the "mostly male hypothesis" (see Frohlich and Chase 2007) in which early blade-like stamens became carpels by the genetic accident of ovules popping up where pollen sacs should have been. Such things do happen, and are reminders that leaves and seed-leaves were originally one and the same.
The third possibility raised by the current phylogentic conclusions is that, instead of simply folding around the ovules, the first carpels formed by an ascidiate growth pattern, i.e. the base of the ancient ovule-bearing structure, or a leaf below it, formed a cup-like base that grew up around a group of ovules (or conceivably a single ovule as in Amborella, but I have already argued agains that in "What's so primitive about Amborella?"). At the same time, the backside of this cup-like structure would develop as a strong backbone, resembling the mid-rib of a leaf, the open top developed a folded structure, and the ovules would come to be placed in two rows opposite the backbone. In terms of genetic and developmental processes, this seems to be a much more complex scenario than simply folding a leaf together. If this is indeed what happened, we need fossil evidence and/or genetic-developmental evidence to confirm it.
If there were a selective pressure for enclosing ancient ovules, the principle of evolution along the lines of least resistance (Stebbins 1974) would clearly favor the easier path of a folding leaf. (see "G. L. Stebbins and the process of adaptive modification" for a full and detailed explanation of this evolutionary principle).
 |
| Though not directly ancestral to the angiosperms, the seed-leaves of the living gymnosperm genus Cycas illustrate the kinds of structures that might have folded together to form the first carpels. Drawing from Asa Gray, 1879. |

Ascidiate carpels most likely evolved among early crown group angiosperms, and presumably evolved for a reason. Most ascidiate carpels, at least those in Amborella and most Austrobaileyales, mature as drupes or berries. These brightly colored fleshy fruits may have been adaptations for improved dispersal by birds in shady forest environments, where these archaic plants survive today.
 |
| A number of gymnosperms, such as this yew (Taxus) has a fruit-like layer that grows up around each seed. Similar features can be found widely among different angiosperms, including magnolids, eudicots and monocots, and may have been present in the earliest angiosperms. Photo by Didier Descouens, posted on Wikipedia. |
In this scenario, the evolution of the ascidiate growth form was an adaptation to embed the ovules more securely within a uniform, sealed wall. The lower tissues could have grown together while leaving the open folded region just at the top. Something similar happened in the evolution of both roses and apples, where tissues of the receptacle were extended up and around the separate carpels.
Now I return to the apparent re-evolution of plicate carpels from ascidiate carpels, as predicted by cladistic analysis. It is odd that simple, leaf-like carpels with marginal rows of ovules would have evolved from the decidedly less leaf-like ascidiate structure, rather than directly from an ancient carpel of essentially the same design. The nature of the suture in modern plicate carpel strongly suggests the joining of opposite edges, and that ovules were attached in rows along those edges.
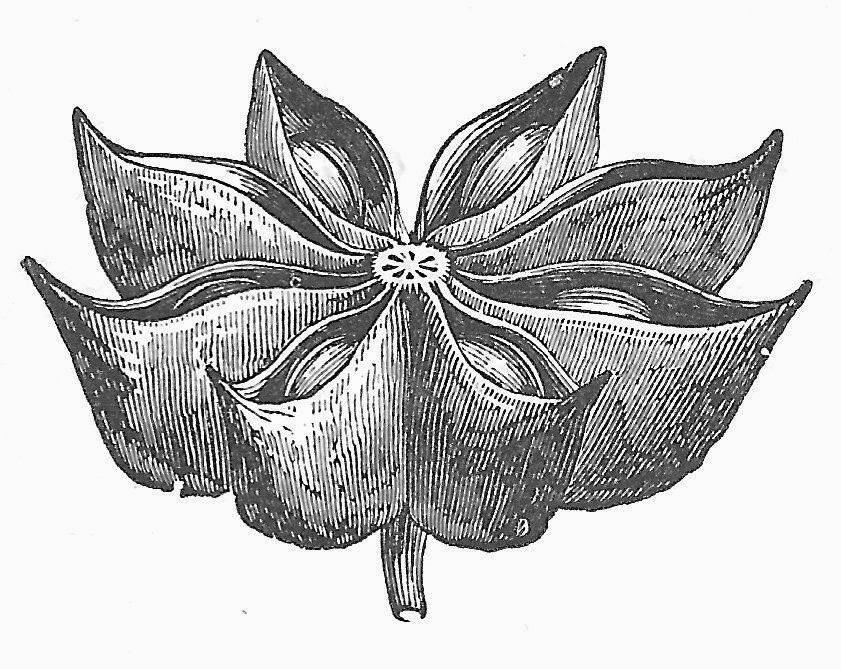 |
| The carpels of Illicium are folded and split open to release seeds, but only one seed is produced per carpel, rather than a row along each margin. They may have evolved from an ascidiate carpel through expansion of the folded stigmatic region. Drawing from Kerner & Oliver, 1895. The Natural History of Plants. |
A simpler adaptive scenario is that a folded carpel with marginal rows of ovules was retained in some ancient crown group angiosperm, and this evolved directly into the more advanced form of plicate carpel with sutures found in the higher angiosperms. The folded nature of the stigmatic region in Amborella may in fact be a remnant of the earliest folded carpels, and the two rows of ovules in Austrobaileya another remnant. So the pieces of the earliest folded carpels are still present among the ANITA grade, and may have been still together in the ancestor of magnolids, monocots, and eudicots.
References:
Bailey, I. W. and G. L. Swamy. 1948. Amborella trichopoda Baill., J. Arnold Arbor. 23:245-254, plus plates.
Endress, P. & J. Doyle. 2009. Reconstructing the ancestral angiosperm flower and its initial specializations. Am. J. Bot. 96(1): 22-66.
Endress, P. & J. Doyle. 2009. Reconstructing the ancestral angiosperm flower and its initial specializations. Am. J. Bot. 96(1): 22-66.
Stebbins,
G. L. 1974. Flowering Plants. Evolution above the species level. Belknap Press of Harvard University
Press. Cambridge, MA.


